С чего начинается процесс видообразования. Видообразование в эволюции — Гипермаркет знаний
Видообразование - процесс возникновения одного или нескольких новых видов в результате изменения системы популяции с генетически открытой на генетически изолированную.
Основные этапы образования вида
1. Географическая изоляция популяции либо глубокое изменение условий среды обитания в части ареала внутри отдельной популяции (биологическая изоляция).
2. Формирование и накопление мутаций внутри популяции, в результате чего изменяется и становится неоднородным генофонд.
3. Сохранение полезных мутаций в ходе естественного отбора.
4. Репродуктивная изоляция, то есть возникновение нового вида.
Прокомментируем данные этапы. Образование новых видов происходит под влиянием изоляции групп особей, иначе говоря, при возникновении препятствий, которые значительно затрудняют свободное скрещивание между особями, принадлежащими как к разным популяциям, так и к одной.
Результатом этого становятся мутации, из которых только полезные мутации закрепляются естественным отбором. В результате невозможности свободного скрещивания постепенно происходит образование нового вида.
Напомним, что факторами видообразования являются все существующие элементарные факторы эволюции, а движущими силами - борьба за существование и естественный отбор.
1. Аллопатрическое видообразование (географическое)
1) Чаще всего новые виды возникают в результате пространственной изоляции популяций, то есть из-за разделения географических ареалов. Такое видообразование носит название аллопатрического или географического.
2) В ходе долгого разобщения единой популяции возникает явление генетической изоляции, которое остается даже в случае, если изоляция прекращается. Аллопатрическое видообразование является достаточно длительным процессом.
3) Новые виды, зародившиеся в результате географического видообразования, часто именуются «географическими расами».
Примеры географического видообразования.
1) Наличие трех подвидов синицы большой: южно-азиатского, евроазиатского и восточно-азиатского.
2) Деление ареала майского ландыша, т. е. формирование сразу нескольких географических рас привело к появлению собственно ландыша майского, ландыша Кейске, ландыша закавказского и др.
3) Наличие двух видов больших чаек: клуши и серебристой чайки в районе Балтийского и Северного морей.
2. Симпатрическое видообразование (экологическое)
Новые виды, образованные с помощью симпатрического (экологического) способа видообразования часто именуют «экологическими расами». Особенностью его является то, что новый вид появляется в пределах популяции с биологической изоляцией.
Примеры экологического видообразования.
1) Сезонные расы погремка большого - в результате вмешательства человека (регулярное скашивание трав) растение не смогло давать семена летом, в результате чего появились подвиды погремка, различающиеся по срокам цветения.
2) Пять различных рас форели в озере Севан (Армения), обитающие на разных глубинах при разных температурах.
3) Двенадцать видов рыб-цихлид в озере Виктория (Восточная Африка), которые значительно различаются по образу жизни, поведению и морфологии.
К симпатрическому видообразованию можно отнести случаи появления новых видов на основе полиплодии и отдаленной гибридизации: например, разные виды картофеля с уникальным набором хромосом (12, 24, 48, 72), хризантем.
Полиплоиды чаще всего более конкурентоспособны и жизнестойки - они могут даже вытеснить родительский вид. Помимо растений, полиплоидия замечена у некоторых видов животных: членистоногих, иглокожих, кольчатых червей и др.
Также в природе может происходить отдаленная гибридизация между видами с последующим удвоением хромосом в геноме. В качестве примера укажем растение рябинокизильник, которые стал гибридом кизильника и рябины. Считается, что порядка 30 процентов всех цветковых появились в результате гибридизации.
Более подробно ознакомиться с темой можно в процессе просмотра видео- и онлайн-лекций автора данных курсов по биологии.
Хочешь сдать экзамен на отлично? Жми сюда -Все новые виды образуются из старых. Происходит это на основе адаптивных преобразований популяций вида или при получении гибридов разных видов.
Вид
Вид можно рассматривать как форму адаптации живой материи к среде обитания и, одновременно, как опорный пункт на бесконечном пути эволюции.
Одним из критериев вида является его генетическая изоляция - нескрещиваемость с другими видами. Если бы её не было, при хаотичном скрещивании организмов происходило бы вымирание потомства и развитие оставалось бы на самом низком уровне.
Пути образования новых видов
Наиболее распространено дивергентное видообразование - расхождение исходного вида на два или несколько новых.
Противоположным ему является путь объединения разных видов в новый на основе гибридизации.
ТОП-4 статьи которые читают вместе с этой
Третьим путём видообразования является филетическое преобразование одного вида в другой. Считается, что таким путём шла эволюция лошади.

Рис. 1. Пути видообразования.
Условно эти три пути можно изобразить:
- А → В, С дивергентное;
- А + В = С гибридогенное;
- А → В филетическое.
Гибридогенное видообразование характерно для растений. Основные способы видообразования животных - дивергентное и филетическое.
Основными формами образования видов являются:
- аллопатрия;
- симпатрия;
- экологическое видообразование.
Явление аллопатрии - ответ на вопрос, является ли географическая изоляция фактором видообразования.
При разделении популяций природными объектами (реки, горы, степи и др.) происходит их обособление, что в дальнейшем приводит и к биологической изоляции (образованию новых видов).
Примерами аллопатрического видообразования является разделение исходного вида ландыш майский на 5 географических рас и возникновение целой системы рас большой синицы.

Рис. 2. Аллопатрическое видообразование у большой синицы.
При симпатрическом видообразовании нет географического разделения между обособляющимися формами. Обычно этот процесс осуществляется на основе полиморфизма вида. Какая-то форма получает преимущество и вытесняет остальные.
Примером является доминирование темноокрашенной формы берёзовой пяденицы над светлоокрашенной в условиях промышленного загрязнения среды.
При экологическом видообразовании вид делится на экологические расы.
Это популяции, которые расходятся по каким-то условиям:
- почвенным;
- климатическим;
- сезонным и т. д.
Постепенно, под действием изолирующих механизмов расы могут обособиться вплоть до генетической (биологической) изоляции.
Изолирующие механизмы предполагают прекращение движения генов между популяциями.
Примером экологического видообразования является процесс разделения севанской форели на 5 сезонных рас.

Рис. 3. Сезонные расы севанской форели.
Симпатрическая и экологическая формы иногда рассматриваются как одна.
Этапы видообразования
По современным представлениям учёных гибридогенное видообразование имеет наиболее быстрые темпы. Иногда достаточно нескольких поколений, чтобы возникший вид закрепился и стал успешно конкурировать с родительским.
Дивергентное растягивается на сотни тысяч и миллионы лет. Оно является основным путём видообразования и имеет в общем виде следующие стадии:
- накопление различий между популяциями;
- появление подвидов;
- накопление различий между подвидами;
- появление биологического типа изоляции между подвидами;
- становление нового вида.
Происходит ли процесс видообразования в настоящее время, и можно ли его наблюдать? Ответ опубликован 6 марта 2008.
Автор вопроса: Алена.
Вопрос задан 2 декабря 2007.
Краткий ответ: процесс видообразования у самостоятельно живущих организмов (не вирусов) в настоящее время происходит со скоростью порядка один новый вид в год на всю биосферу, но наблюдать этот процесс невозможно в силу недостаточной разрешающей способности существующей техники таких измерений.
Подобная ситуация часто встречается в науке: из твердо установленных эмпирических данных и теорий достоверно известно, что процесс существует, но прямое наблюдение его пока невозможно вследствие недостаточной разрешающей способности техники измерений.
1. Эволюция происходит
Установлено, что время существования вида не зависит от размеров особей и численности популяций вида и одинаково для одноклеточных видов с огромными численностями популяций и для многоклеточных видов млекопитающих, птиц и др., численности популяций которых на много порядков величины меньше.
Откуда известно, что эволюция происходит с указанной выше скоростью, один новый вид на биосферу в год? Это известно из двух твердо установленных фактов:
1) существования абсолютной шкалы времени по данным о распаде радиоактивных изотопов,
2) палеоданных, т.е. раскопок останков вымерших видов.
Никаких других свидетельств существования эволюции нет. Согласно палеоданным, все виды оставались неизменными (что и позволяет причислять останки к определенному виду) в течение времени в среднем порядка T
= 3 x 10 6 (3 миллиона) лет. Затем виды вымирали. Установлено, что это время не зависит от размеров особей и численности популяций вида и одинаково для одноклеточных видов с огромными численностями популяций и для многоклеточных видов млекопитающих, птиц и др., численности популяций которых на много порядков величины меньше. Количество видов, населявших биосферу в каждый момент времени, несмотря на значительные колебания, не меняло порядка своей величины N
и составляет сейчас от 3 до 10 миллионов видов, т.е. появление новых видов должно было сопровождаться вымиранием старых. Скорость эволюции, следовательно, можно оценить отношением N
/T
~ один-три вида в год.
2. Видообразование и внутривидовая изменчивость не связаны между собой
Эволюция происходит путем изменения генетической информации вида, т.е. путем изменения структуры ДНК, образующей линейную последовательность генетических букв (нуклеотидных пар), которую называют геномом вида. Относительное различие геномов двух родственных видов примерно одинаково у любой пары родственных видов и не зависит от размера тела особей и численности популяций этих видов. Частота мутаций (число наблюдаемых замен нуклеотидных пар в геноме, происходящих в единицу времени) определяется молекулярной структурой ДНК и, следовательно, одинакова у всех особей вида. Поэтому, чем больше численность популяции вида, тем больше мутационных замен накапливается в единицу времени во всей популяции. Если бы внутрипопуляционные замены определяли скорость эволюции, т.е. какая-нибудь из накопившихся в популяции мутационных замен приводила бы к образованию нового вида, то скорость эволюции была бы пропорциональной численности особей популяции, что противоречит палеоданным. Следовательно, эволюция не может происходить за счет наблюдаемых случайных мутаций (изменений) геномов особей, возникающих внутри популяции отдельного вида.
Внутривидовая изменчивость ограничена стабилизирующим отбором. Если количество ошибок в генетической информации особи превосходит порог разрешающей способности конкурентного взаимодействия особей внутри популяции, то такие особи воспринимаются остальными особями как уродливые и удаляются из популяции. Особи с числом ошибок, меньшим этого порога, воспринимаются как нормальные. В силу случайности ошибок, в разных популяциях вида они накапливаются в разных участках генома. Ошибки могут случайно фиксироваться у всех особей популяции и длительное время передаваться по наследству. Это приводит к подвидовым и расовым отличиям особей различных популяций. Однако несмотря на то, что в геномах разных рас и подвидов зафиксированы разные ошибки, все расы и подвиды остаются одинаково удаленными от нормального генома на величину, меньшую порога чувствительности стабилизирующего отбора, и поэтому являются генетически равноценными. Мутационный процесс и стабилизирующий отбор приводят к появлению ошибок в одних местах генома и исчезновению равного количества ошибок в других местах генома. Подобный дрейф мутационных замен не увеличивает отклонения от нормального генома вида и не меняет основных видовых характеристик особи, позволяющих однозначно определять принадлежность особи к определенному существующему или вымершему виду.
Искусственный отбор, в отличие от генетического дрейфа в естественных популяциях, сводится к направленному отбору уродливых особей вида, используемых человеком для своих целей.
Таким образом, все наблюдаемые внутривидовые мутации, приводящие к изменению особей вида, включая любые варианты искусственного отбора, гибридизацию, генно-инженерную модификацию, не соответствуют образованию новых видов.
3. Внешний фактор процесса видообразования - РНК вирусы
Следовательно, эволюция происходит за счет внедрения в геномы особей вида внешних фрагментов генетической функционально осмысленной информации. Источник внешней генетической информации должен посылать эту информацию во все виды биосферы с одинаковой скоростью, через одинаковые промежутки времени порядка 3 миллионов лет. Следовательно, этот источник должен иметь единую универсальную природу для всей биосферы.
Какова природа этого универсального источника внешней генетической информации?
Для построения белков и активизации всех остальных процессов в клетке информация ДНК (главного хранилища) переписывается на большое число коротких макромолекул РНК, которые служат матрицами для синтеза белка. В ДНК встроена программа репарации (починки) мутационных ошибок, возникающих при копировании молекул ДНК при делении клетки. Это уменьшает частоту мутаций в ДНК в миллион раз по сравнению с частотой мутации в РНК, для которой программы репарации нет.
В природе существуют РНК-вирусы (ретровирусы), чья видовая генетическая информация записана не на молекулах ДНК, а на макромолекуле РНК. Поэтому частота мутаций у таких вирусов в миллион раз больше, чем у любых других организмов, хранящих свою генетическую информацию в ДНК. Вирусы мертвы вне клетки. Активными вирусы становятся только внутри клетки хозяина. Вирус иммунодефицита человека (ВИЧ), вызывающий СПИД, обнаруженный в конце прошлого века, относится к ретровирусам. Есть все основания полагать, что этот вирус не существовал ранее, и его возникновение есть наблюдаемый пример эволюционного процесса (но не у живых организмов, а у вирусов). Это пример эволюционного появления новой программы разрушения самостоятельно живущих организмов существующего вида (Homo sapiens). ВИЧ поражает нормальную генетическую программу человека.
РНК вирусы изменяются и эволюционируют случайно. Они не могут приготавливать прогрессивную информацию, направленную на эволюцию одного выделенного вида биосферы. Поэтому вероятность появления новой прогрессивной информации для любого вида биосферы одинакова. Именно это объясняет одинаковую скорость эволюции для всех существующих видов биосферы независимо от их размера тела и численности популяции особей. Это исчерпывает наш ответ на поставленный вопрос.
4. Традиционные взгляды на эволюцию; креационисты и эволюционисты
Рассмотрим также, какой ответ дается на поставленный вопрос в профессиональных кругах эволюционистов-биологов. Краткий ответ такой же, что и был дан нами вначале, а именно: эволюция происходит и сейчас со скоростью порядка одного вида в год на всю биосферу, но заметить это пока невозможно. Однако в отношении причин, вызывающих эволюцию, не существует единого мнения.
До сих пор в профессиональных кругах эволюционистов ведутся споры о том, является ли эволюция непрерывной или скачкообразной. В действительности, как хорошо известно в физике, непрерывность или дискретность связана с рассмотрением пространственного или временного масштаба процесса. Например, газ имеет непрерывное давление и температуру на масштабах, намного превосходящих расстояния между молекулами, но все характеристики газа становятся дискретными на масштабах порядка или меньше межмолекулярных расстояний. На масштабах времени, много больших времени существования видов, и масштабах пространства, включающих всю макромолекулу ДНК, эволюция может рассматриваться как непрерывная. Очевидно, что геномы шимпанзе и человека имеют фундаментальные различия в генетической информации, но где в геноме распределены эти различия и как они выглядят на молекулярном уровне, пока установить не удается. Наблюдаемые количественные различия между нуклеотидными парами (генетическими буквами) геномов двух случайно выбранных особей человека или геномов двух случайно выбранных особей шимпанзе не отличаются от соответствующих различий между человеком и шимпанзе , т.е. дискретность эволюции пока не обнаруживается экспериментально на молекулярном уровне.
Все виды биосферы, включая все близкородственные виды, дискретны и жестко разделены между собой. Не наблюдается непрерывных переходных форм от одного вида к другому как для всех существующих сейчас видов, так и для всех вымерших видов в палеоданных. Этот факт лежит в основе существования видов: были бы непрерывные переходные формы, не было бы видов вообще.
Процесс возникновения видов (т.е. процесс превращения старого вида в новый) не обнаруживается по палеоданным. Известно лишь, что этот процесс занимает намного меньше среднего времени существования вида (3 x 10 6 лет) и не превосходит десяти тысяч лет. Т.е. вид возникает практически мгновенно, время возникновения вида меньше долей процента времени существования вида. Это прекрасно согласуется с описанным выше механизмом "прогрессивного заражения" вида фрагментами генома, приготовленными вирусами. Дискретность эволюционного процесса на масштабах времени порядка 10 4 лет << T ~ 3 x 10 6 лет и непрерывность эволюции на масштабах времени, превышающих T ~ 3 x 10 6 лет, являются важнейшими наблюдаемыми характеристиками эволюции.
Непрерывность эволюции приписывается дарвиновской теории естественного отбора. Отказ от непрерывности эволюции, следовательно, означает отказ от дарвиновской теории. Факт дискретности видов берется на вооружение так называемыми креационистами, связывающими эволюцию с внешней внебиосферной силой. Креационисты в действительности не утверждают ничего нового по сравнению с описанным нами выше фактом, что виды возникают под воздействием внешних по отношению к виду агентов, а не под воздействием внутривидовых изменений. Сторонники "дарвиновской" непрерывной теории эволюции (которая в свете современных данных стала носить название "неодарвинизма") вынуждают сторонников скачкообразной эволюции, основанной на палеоданных, отказываться от своих воззрений. (Ситуация мало отличается от времен Галилея.)
Теория естественного отбора Дарвина сделала ненужной использование внешней силы ("творца", существующего вне биосферы) для объяснения происхождения видов. Стало ясно, что эволюция может происходить за счет процессов внутри самой жизни, т.е. внутри биосферы. И это утверждение, являющееся огромной заслугой Дарвина, является правильным и непротиворечивым. В то же время Дарвин полагал, что процесс естественного видообразования в природе происходит в результате наблюдаемой внутривидовой изменчивости и аналогичен процессу искусственного отбора. Но во времена Дарвина не была открыта абсолютная шкала времени. Не была известна обсуждавшаяся выше независимость продолжительности существования вида от размеров особей и численности популяции. Дарвин не мог знать об описанных выше фундаментальных различиях между внутривидовой изменчивостью, искусственным отбором и естественным видообразованием. Это было выяснено позже. Поэтому основная концепция Дарвина остается правильной, она включает неизвестную Дарвину дискретность видообразования на масштабах времени, много меньших миллиона лет, и предполагаемую Дарвиным непрерывность эволюции на масштабах времени, много больших миллиона лет. Неверным оказалось лишь отождествление внутривидовых изменений и искусственного отбора с процессом естественного видообразования. Именно оно в конечном счете делает традиционную картину эволюции уязвимой по отношению к аргументам креационистов.
Следует отметить, что Линней, сформулировавший за сто лет до Дарвина принципы систематики видов, фактически утверждал, что виды из одного и того же рода различаются между собой меньше, чем виды из разных семейств, и, тем более, из разных отрядов и классов. Линней писал, что виды сотворены внешней силой, что верно (виды не возникают в результате внутривидовых изменений), и остаются неизменными, что также верно (виды не меняются в течение миллионов лет). Линней не мог не понимать, что виды из одного и того же рода возникли из одного предшествовавшего вида, т.е. их "сотворение" было скоррелированным (иначе эти родственные виды не были бы столь похожи друг на друга). Существование эволюционного процесса однозначно вытекало из систематики Линнея, независимо от Дарвина. Если бы виды создавались внешней к биосфере силой ("творцом"), то никакие виды не были бы похожими и все виды одинаково случайно отличались бы друг от друга. Не было бы систематики, открытой Линнеем. По-видимому, прекрасно понимая это и признавая сотворение неменяющихся со временем видов внешней силой ("творцом"), Линней сумел добиться прижизненного признания своих трудов, искусно избавившись от гонений и травли господствующего в те времена общественного мнения, близкого к мнению современных креационистов.) Лишь теперь стало ясно, кто может быть кандидатом на внешнюю силу Линнея - это РНК вирусы. Эта внешняя по отношению к эволюционирующему виду сила находится внутри биосферы, внутри самой жизни, не требует привлечения понятия "творца" и не противоречит основному положению концепции Дарвина.
- Эволюционная теория атакована: дискуссия, в которой недостает количественных аргументов? (о том, убедительны ли ответы эволюционистов креационистам) (на англ. яз.)
Видообразование - это сложнейший эволюционный процесс возникновения нового вида. Новый вид прерывает связи с родительским видом и превращается в обособленную совокупность организмов. Скрещивание особей нового и старого видов становится невозможным.
Механизм видообразования. Представление о механизмах видообразования впервые было высказано Ч. Дарвином. Он исходил из того, что внутривидовая борьба за существование и вытекающий из нее естественный отбор служат главной причиной расхождения популяций, заставляют виды максимально широко и разнообразно использовать природные условия. По мнению Ч. Дарвина, в пределах одного вида часто возникают популяции, которые приспосабливаются к разным условиям обитания: влажным или сухим, равнинным или горным местообитаниям, потреблению определенной пищи и т. д. Именно благодаря этому виды с наибольшей полнотой используют ресурсы среды своего обитания. Следовательно, естественный отбор благоприятствует все более полному использованию разнообразия условий существования. Это вызывает расхождение популяций в пределах вида по морфологическим, физиологическим и биохимическим признакам.
Географическая и экологическая изоляция, как уже отмечалось, - эволюционные механизмы, резко усиливающие процесс расхождения популяций вследствие ослабления и даже полного прекращения обмена генами между ними.
Рассмотрим случай, когда видовой ареал оказывается расчлененным на две части, например в результате возникновения пролива, отделившего полуостров от материка. Следствием такого расчленения ареала является разбивка вида на две не скрещивающиеся друг с другом популяции. Единый генофонд вида в результате оказывается разделенным на две изолированные части. (Генофондом называют совокупность всех генов, содержащихся в популяции или виде.)
Изначальное различие между генофондами обособившихся частей вида в ходе дальнейшей эволюции неизбежно усиливается. В качестве важной причины этого выступает мутационный процесс. В одной из двух разделившихся популяций возникают одни мутации, а в другой - иные. Поскольку условия обитания в разных «изоля-тах» могут существенно отличаться, то и естественный отбор также будет действовать в разных направлениях. Дрейф генов также вносит важный вклад в дальнейшее расхождение генофондов обособившихся частей вида. Процесс расхождения популяций, продолжаясь длительное время, приводит к глубоким изменениям генофондов популяций и многих свойств организмов: размеров, окраски, плодовитости, предпочтения определенных мест обитания и т. д. Возникают новые признаки и адаптации, появляются отличия в строении и даже количестве хромосом.
Вследствие этого особи прежде родственных популяций утрачивают способность скрещиваться между собой. Возникают новые виды. В результате видообразования появляется вид или несколько видов, которые никогда прежде не существовали. Они обладают только им присущими свойствами.
Выделяют два основных способа видообразования: географическое и экологическое. Различие между ними состоит в том, какой тип изоляции послужил исходным для расхождения популяций. Сущность процесса видообразования одинакова для обоих способов. Она, как уже было отмечено, заключается в коренной перестройке свойств родительского вида и в формировании свойств нового вида.
Большая часть вновь возникающих видов бракуется. Только наиболее успешно осваивающие новые экологические условия, т. е. наиболее приспособленные, виды сохраняются и идут по пути дальнейшего эволюционного развития.
- Какую роль в образовании новых видов играют изоляция, дрейф генов, популяционные волны?
- Опишите два основных способа видообразования.
- Происходит ли процесс видообразования в наше время и можно ли его наблюдать?
- Объясните, как, по вашему мнению, могли образоваться из одного родоначального вида два вида зайцев - беляк и русак.
- Может ли, по вашему мнению, деятельность человека повлиять на образование новых видов? Ответ обоснуйте.
В территориальном аспекте в пространстве новый вид может возникнуть из одной или группы смежных популяций, расположенных на периферии исходного вида. Такое видообразование называется аллопатрическим (от греч. alios - иной, patris - родина). В других случаях новый вид может возникнуть внутри ареала исходного вида, как бы внутри вида; этот путь видообразования называется симпатрическим (от греч. sym - вместе, patris - родина). В филогенетическом аспекте (во времени) новый вид может возникнуть посредством постепенного изменения одного и того же вида во времени, без какой-либо дивергенции исходных групп. Такое видообразование называется филетическим.
Новый вид может возникнуть путем разделения единого предкового вида (дивергентное видообразование). Наконец, новый вид может возникнуть в результате гибридизации двух уже существующих видов - гибридогенное видообразование. Кратко опишем эти основные пути видообразования.
Аллопатрическое видообразование (называемое иногда географическим) иллюстрируется приведенными выше примерами возникновения вида у больших чаек и в группе австралийских мухоловок. При аллопатрическом видообразовании новые виды могут возникнуть путем фрагментации, распадения ареала широко распространенного родительского вида. Примером такого процесса может быть возникновение видов ландыша (см. гл. 6). Другой способ аллопатрического видообразования - видообразование при расселении исходного вида, в процессе которого все более удаленные от центра расселения периферийные популяции и их группы, интенсивно преобразуясь в новых условиях, становятся родоначальниками видов. Примеры, аналогичные видообразованию в группе больших чаек, известны и для других птиц, некоторых рептилий, .
В основе аллопатрического видообразования лежат те или иные формы пространственной изоляции, и этот путь видообразования всегда сравнительно медленный, происходящий на протяжении сотен тысяч поколений. Именно за такие длительные промежутки времени в изолированных частях населения вида вырабатываются те биологические особенности, которые приводят к репродуктивной самостоятельности даже при нарушении первичной изолирующей преграды. Аллопатрическое видообразование всегда связано с историей формирования видового ареала.
Симпатрическое видообразование. При видообразовании симпатрическим путем новый вид возникает внутри ареала исходного вида.
Первый способ симпатрического видообразования - возникновение новых видов при быстром изменении кариотипа, например при автополиплоидии. Известны группы близких видов (обычно ) с кратными числами хромосом (см. рис. 6.28). Так, например, в роде хризантем (Chrysanthemum) все формы имеют число хромосом, кратное 9, 18, 27, 36, 45, ..., 90. В родах табака (Nicotiana) и картофеля (Solanum) основное, исходное, число хромосом равно 12, но имеются формы с 24, 48, 72 хромосомами. В таких случаях можно предположить, что видообразование шло путем - посредством удвоения, утроения, учетверения и т. д. основного набора хромосом предковых видов. Процессы полиплоидизации хорошо воспроизводятся в эксперименте посредством задержки расхождения хромосом в мейозе в результате воздействия, например, колхицином. Известно, что полиплоиды могут возникать и в природных условиях. Возникшие полиплоидные особи могут давать жизнеспособное потомство лишь при скрещивании с особями, несущими то же число хромосом (или при самоопылении). В течение немногих в том случае, если полиплоидные формы успешно проходят «контроль» и оказываются лучше исходной , они могут распространиться и сосуществовать совместно с породившим их видом (рис. 13.4) или, что бывает чаще, просто вытеснить его.
Рис. 13.4. Пример возникновения репродуктивной изоляции при полиплоидизации: обитающий по всему Индостану тетраплоидный вид растений Dicanthium annulatum, несомненно, возник из диплоидной предковой формы, ныне занимающей небольшой дизъюнктный ареал (по Н. Россу, 1962)
Полиплоидные формы, как правило, крупнее и способны существовать в более суровых физико-географических условиях. Именно поэтому в высокогорьях и в Арктике число полиплоидных видов растений резко увеличено (рис. 13.5). Среди полиплоидия при видообразовании играет несравненно меньшую роль, чем у растений, и во всех случаях связана с способом размножения (например, у , членистоногих, аннелид и других беспозвоночных).
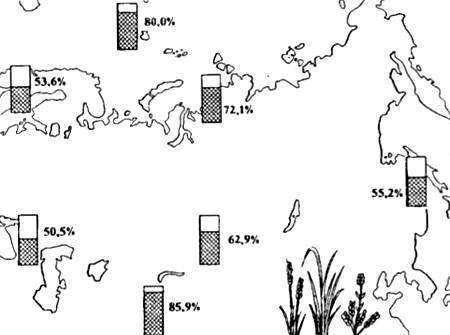
Рис. 13.5. Распространение полиплоидных видов цветковых (в процентах к общему числу видов флоры) в разных частях Евразии (по данным разных авторов из Н.В. Тимофеева-Ресовского и др., 1977)
Второй способ симпатрического видообразования - путем гибридизации с последующим удвоением числа хромосом - аллополиплоидия (см. ниже).
Наконец, последним, достаточно изученным способом симпатрического видообразования является возникновение новых форм в результате сезонной изоляции. Известно существование ярко выраженных сезонных рас у растений, например у погремка Alectorolophus major (см. гл. 10), раннецветущие и позднецветущие формы которого полностью репродуктивно изолированы друг от друга, и, если условие отбора сохраняется, лишь вопрос времени - когда эти формы приобретут ранг новых видов. Аналогично положение с яровыми и озимыми расами проходных рыб; возможно, что эти формы уже являются разными видами, очень схожими морфологически, но изолированными генетически (виды-двойники).
Особенность симпатрического пути видообразования - возникновение новых видов, морфофизиологически близких к исходному виду. Так, при полиплоидии увеличиваются размеры, но общий облик растений сохраняется, как правило, неизменным; при хромосомных перестройках наблюдается та же картина; при экологической (сезонной) изоляции возникающие формы также обычно оказываются морфологически слабо различимыми. Лишь в случае гибридогенного возникновения видов появляется новая видовая форма, отличная от каждого из родительских видов (но также имеющая признаки, характерные для двух исходных видов).
Филетическое видообразование. При филетическом видообразовании вид, весь в целом изменяясь в чреде поколений, превращается в новый вид, который можно выделить, сопоставляя морфологические характеристики этих групп.
Филетическое видообразование включает стазигенез - развитие вида во времени с постепенным изменением одной и той же экологической ниши, и анагенез - развитие вида с приобретением каких-то новых принципиальных приспособлений, позволяющих ему образовать совершенно новую, более широкую экологическую нишу. Примером стазигенеза может быть развитие основного ствола верхнеплиоценовых моллюсков рода Giraulus (см. рис. 6.5).

Рис. 13.6. Пример филетического видообразования в ряду ископаемых европейских слонов (Elephas planifrons - Е. meridionalis) по ламеллярному индексу (количеству эмали на зубах) (из В. Гранта, 1980)
Ясно, что в этом случае возможно лишь сопоставление морфологических характеристик, так как изучать результаты филетической эволюции можно лишь с привлечением палеонтологического материала (рис. 13.6). При этом всегда остается возможность того, что на каком-то этапе эволюции от единого филетического ствола могли дивергировать другие группы и филетическое видообразование могло оказаться на самом деле дивергентным. Поэтому в «чистом виде» филетическая эволюция, видимо, возможна лишь как идеализированное и упрощенное отражение эволюционного процесса на одном из отрезков жизни вида (фратрии).
Заметим, что границы между отдельными видами в филетическом ряду форм провести невозможно - она всегда будет условной (см. гл. 12).
Дивергентное видообразование (кладогенез). Ч. Дарвин считал этот тип видообразования самым распространенным (единственный рисунок в «Происхождении видов» посвящен именно этому типу видообразования). Примерами этого типа видообразования - возникновение новых видов в результате разделения единой предковой формы - являются возникновение нескольких видов сигов вокруг (см. рис. 6.10) и дивергенция дарвиновых вьюрков на Галапагосах (см. рис. 6.12) и эволюция североамериканских дрозофил группы pseudoobscura - persimilis (см. рис. 6.27).
Гибридогенное видообразование (синтезогенез или сингенез). Этот тип видообразования обычен у растений: по некоторым подсчетам, более 50% видов растений представляют собой гибридогенные формы - аллополиплоиды. Укажем лишь на некоторые. Культурная слива (Prunus domestica) с 2n = 48 возникла путем гибридизации терна (P. spinosa, 2n = 32) с алычой (P. divaricata, 2n= 16) с последующим удвоением числа хромосом. Некоторые виды пикульника, малины, табака, брюквы, полыни, ириса и других растений - такие же аллополиплоиды гибридогенного происхождения.
Интересен случай возникновения нового симпатрического вида у Spartina townsendii (2л = 120) на основе гибридизации с последующим удвоением числа хромосом местного английского вида S. stricta (2n = 50) и завезенной в 70-х годах XIX в. из Северной Америки S. alternifolia (2n = 70). Сейчас ареал этого вида интенсивно расширяется за счет сокращения ареала местного европейского вида. Другой пример гибридогенного вида, возникшего симпатрически,- рябинокизильник (Sorbocotaneaster), сочетающий признаки рябины и кизильника и распространенный в середине 50-х годов в лесах южной Якутии по берегам среднего течения р. Алдан (К.М. Завадский). В результате гибридогенного видообразования особенно часто могут образовываться комплексы видов (или так называемых полувидов), связанных между собой гибридизацией,- сингамеоны (В. Грант). В случае таких гибридных комплексов иногда бывает трудно обнаружить четкие границы между отдельными видами, хотя виды как устойчивые генетические системы выделяются вполне определенно.
Все четыре основные формы видообразования во времени схематично показаны на рис. 13.7.

Рис. 13.7. Основные формы филетического (во времени) видообразования (из Н.Н. Воронцова, 2001)